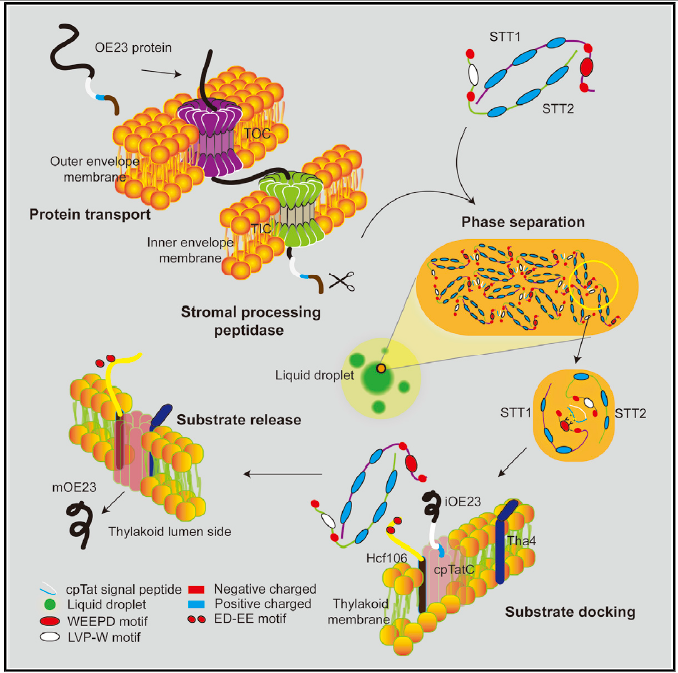
STTs介导的相分离驱动叶绿体内蛋白分选机制
2020年3月12日,作物逆境适应与改良国家重点实验室张立新教授课题组及合作者在光合作用研究领域取得重要进展,在国际上首次提出并阐明了相分离驱动叶绿体内蛋白分选的新机制,推动了蛋白转运机理的进一步深入,揭示了相分离的重要生理意义,而且对于探讨叶绿体的生物发生、光合器官的建成和功能调节以及真核生物的起源和进化等都具有重要的意义。该研究成果以“Liquid-liquid phase transition drives intra-chloroplast cargo sorting”为题,以全文形式在《Cell》杂志在线发表(DOI:10.1016/j.cell.2020.02.045)。张立新教授为本文通讯作者,中国科学院植物研究所是第一作者单位,河南大学是通讯作者单位。
光合作用是地球上最大规模地利用太阳能,把二氧化碳和水合成为有机物,并放出氧气的过程。叶绿体是植物光合作用场所。叶绿体是由光合细菌共生演变而来的,在光合作用及其他多种重要生理过程中发挥着关键性的作用。叶绿体具有半自主性,95%叶绿体蛋白是由核基因编码的,在胞质中合成为前体后,通过叶绿体外被膜和内被膜上的转运通道将蛋白质转入叶绿体的不同区域才能使叶绿体行使光合功能。然而一直以来科学家们对于核编码叶绿体蛋白在跨过叶绿体被膜之后如何被分选、穿过拥挤的基质空间、并精确地靶定到特异性类囊体膜复合物的分子机制依然不清楚。该研究发现了位于叶绿体基质的关键蛋白转运分选因子STT1与STT2,阐明了双精氨酸依赖转运途径的底物识别、分选以及转运靶定到双精氨酸依赖转运途径的分子机制。STT1与STT2蛋白都包含N端的IDR (intrinsically disordered region)结构域与C端的ankyrin repeat结构域。STT1与STT2能够依赖于ankyrin repeat结构域的相互作用形成一个椭圆球状异源二聚体结构,而STT1与STT2的IDR区域分别含有保守的WEEPD基序与LVP-W基序,负责识别底物信号肽的双精氨酸(RR)基序与疏水结构域(H domain)。研究表明底物结合后会激活STT复合物进一步组装相分离形成浓缩的液滴。STT-底物相分离液滴协助底物穿过叶绿体基质从而靶定到类囊体膜。而Hcf106能够抑制STT的相分离从而释放底物,完成底物的正确运输与装配。这种方式可能既能确保底物折叠,同时保持底物信号肽的活性便于被下游Tatc/Hcf106通道识别。阻碍STT-Hcf106结合会阻断Tat底物的运输,影响植物光合作用从而导致植物致死的表型。该研究首次发现了相分离调控叶绿体蛋白转运,从而调控叶绿体生成和光合作用。
该研究揭示了调节细胞器生物发生蛋白转运相分离的保守机制,为研究细胞如何精确调控生理过程拓展了新思路。该研究工作受到国家自然科学基金委、科技部、河南大学等项目的资助。